- Research
- Open access
- Published:
Characterization of acetic acid-detoxifying Escherichia coli evolved under phosphate starvation conditions
Microbial Cell Factories volume 15, Article number: 42 (2016)
Abstract
Background
During prolonged incubation of Escherichia coli K-12 in batch culture under aerobic, phosphate (Pi) starvation conditions, excess glucose is converted into acetic acid, which may trigger cell death. Following serial cultures, we isolated five evolved strains in two populations that survived prolonged incubation.
Methods
We sequenced the genomes of the ancestral and evolved strains, and determined the effects of the genetic changes, tested alone and in combination, on characteristic phenotypes in pure and in mixed cultures.
Results
Evolved strains used two main strategies: (1) the constitutive expression of the Trk- and Kdp-dependent K+ transport systems, and (2) the inactivation of the ArcA global regulator. Both processes helped to maintain a residual activity of the tricarboxylic acid cycle, which decreased the production of acetic acid and eventually allowed its re-consumption. Evolved strains acquired a few additional genetic changes besides the trkH, kdpD and arcA mutations, which might increase the scavenging of organophosphates (phnE +, lapB, and rseP) and the resistance to oxidative (rsxC) and acetic acid stresses (e14−/icd +).
Conclusions
Evolved strains rapidly acquired mutations (phnE + lapB rpoS trkH and phnE + rseP kdpD) that were globally beneficial to growth on glucose and organophosphates, but detrimental to long-term viability. The spread of these mutant strains might give the ancestral strain time to accumulate up to five genetic changes (phnE + arcA rsxC crfC e14−/icd +), which allowed growth on glucose and organophosphates, and provided a long-term survival. The latter strain, which expressed several mechanisms of protection against endogenous and exogenous stresses, might provide a platform for producing toxic recombinant proteins and chemicals during prolonged incubation under aerobic, Pi starvation conditions.
Background
In nature, bacteria are frequently starved for essential nutrients such as phosphate (HPO4 2−, Pi) [1, 2]. When the levels of Pi in the medium decrease below 4 μM, Escherichia coli induces the PhoBR regulon, which helps to scavenge low levels of Pi and secondary sources of Pi such as organophosphates and phosphonates [3]. When the cells cannot find a sufficient source of Pi to maintain growth, they enter the so-called stationary phase [4]. The E. coli K-12 strain MG1655 incubated in batch culture under aerobic, Pi-limiting conditions (0.1 mM K2HPO4 and 40 mM glucose) enters stationary phase after 10 h of incubation when Pi levels drop below 1 μM [5].
At the approach of the stationary phase, the cells steadily accumulate the RpoS (σs) sigma factor [6, 7]. The σs-RNA polymerase holoenzyme poorly transcribes “growth genes” (e.g. genes encoding the succinate dehydrogenase of the tricarboxylic acid cycle) and preferentially transcribes “defense genes” required for the protection of non-growing cells against endogenous stresses such as oxidative stress (e.g. pdhR, poxB, and sodC) and acetic acid stress (e.g. gadB) [4, 8–10]. For instance, cells incubated under Pi starvation conditions continue to metabolize glucose at a reduced rate, but eventually redirect the metabolic flux from the pyruvate dehydrogenase towards the RpoS-dependent pyruvate oxidase, PoxB, which directly converts pyruvate into acetic acid. PoxB (pyruvate:Q reductase), in contrast to the pyruvate dehydrogenase, does not use NAD+ as a cofactor, which prevents the adventitious production of O .−2 and H2O2 by NADH dehydrogenases in the aerobic respiratory chain [4]. Whereas the activity of PoxB protects Pi-starved cells against oxidative stress at the entry into stationary phase, this activity can eventually cause the accumulation of high levels of acetic acid, which decrease the internal pH (pHi), stop metabolism and trigger cell death [4, 10]. Death of Pi-starved cells can be alleviated by the addition of glutamate into the medium, which allows the RpoS-dependent GadB acid resistance system to neutralize acetic acid [5, 10] (Fig. 1).

Schematic illustration of the relationship between pyruvate metabolism and the aerobic respiratory chain. AceCoA acetyl CoA, BCAA branched-chain amino acids, CIT citrate, CS citrate synthase (GltA), FAD flavin adenine dinucleotide, FUM fumarate, GABA γ-aminobutyrate, Glc glucose, Glg glycogen, GLX glyoxylate, GLU glutamate, G6P glucose 6-phosphate, ICT isocitrate, IDH isocitrate dehydrogenase (Icd), IM inner membrane, KG α-ketoglutarate, KGDH α-ketoglutarate dehydrogenase, MAL malate, MDH malate dehydrogenase, NDH NADH dehydrogenase, OA oxaloacetate, PDH pyruvate dehydrogenase, pHi internal pH, PEP phosphoenolpyruvate, PYR pyruvate, Q ubiquinone, SDH succinate dehydrogenase, SUC succinate, SucCoA succinyl CoA; TCA, tricarboxylic acid
Whereas E. coli is normally doomed to die when glutamate is not added in Pi-limiting medium, evolved strains can sweep populations after generally three serial batch cultures. Variants that survive prolonged incubation exhibit different mutant phenotypes refereed to as RpoS− (colonies grown on LB medium produce low levels of glycogen and exhibit a reduced catalase activity) and Glg− (colonies produce reduced levels of glycogen) [1]. From one evolved population, we isolated three strains exhibiting RpoS− (ENZ1901), Glg− (ENZ1902), and Glg+ (ENZ1903) phenotypes. In a parallel-evolved population, which contained a majority of Glg− mutants and eventually Glg+ cells, we isolated two Glg+ evolved strains (ENZ1904 and ENZ1905) [1].
Compared to the ancestral strain, the evolved strain ENZ1901 (RpoS−) harbors a single-nucleotide deletion (rpoS-ΔG214) that inactivates RpoS, and an 8-bp deletion in phnE that activates the PhnE permease [1]. The lack of RpoS activity allows a substantial activity of the tricarboxylic acid cycle in Pi-starved cells, which decreases the production of acetic acid and allows its re-consumption (Ace+ phenotype) [1, 5, 11, 12]. The PhnE permease can scavenge organophosphates excreted into the medium [1]. The combination of the phnE and rpoS evolved alleles triggers a novel growth-under-Pi-starvation (GPS) phenotype, which refers to the ability of a mutant strain—incubated for 1 day in Pi-limiting medium and added in minority into a 1-day-old culture of the ancestral strain in Pi-limiting medium—to resume growth between days 1 and 4 of incubation by using primarily organophosphates released into the mixed culture by the ancestral strain [1]. The GPS phenotype may account at least in part for the spread of the evolved strain ENZ1901 in populations starved for Pi.
If the presence of an rpoS-null mutation may help to explain the behavior of the evolved strain ENZ1901 (RpoS−), the majority of the evolved strains did not exhibit an RpoS-negative phenotype [1]. This prompted us to determine the complete genotypes, the metabolic patterns and the growth capacities in Pi-limiting medium of the evolved and reconstructed mutant strains. We show here that mutants could overtake populations and detoxify acetic acid by using two novel strategies: (1) K+ accumulation (triggered by trkH or kdpD mutations) combined with changes in outer membrane permeability (lapB or rseP mutations), and (2) inactivation of ArcA activity (arcA mutation) combined with an increased resistance to oxidative stress (rsxC mutation) and to acetic acid stress (crfC and e14− / icd + mutations).
Results
Genotype of the evolved strains
Whole-genome sequencing of the evolved strains ENZ1901-ENZ1905 and of the ancestral strain ENZ535 revealed limited changes compared to the MG1655 reference sequence (F− λ− rfb-50 rph-1 ilvG phnE ylbE icd::e14) (ftp://ncbi.nlm.nih.gov/blast/executables/blast+/LATEST/) (Table 1; Fig. 2):

Genetic map of the evolved strains ENZ1901, ENZ1902, and ENZ1903
-
1.
In the ancestral strain ENZ535, the sequence of the ylbE gene differed from the reference sequence by a single-nucleotide polymorphism and a single-nucleotide insertion that removed a stop codon. The so-called ylbE + allele in ENZ535 is also present in the MG1655 strain referred to as ATCC47076 [17]. The function of YlbE is not known.
-
2.
In the five evolved strains (PhnE-expressing), the sequence of the phnE gene differed from the reference sequence by a deletion of 8 bp, which removed a frameshift inactivating phnE [1].
-
3.
The evolved strains ENZ1904 and ENZ1905 (Glg+) acquired no others mutations besides the 8-bp deletion in phnE (phnE +).
-
4.
The evolved strains ENZ1901 (RpoS−), ENZ1902 (Glg−), and ENZ1903 (Glg+) harbored unique genetic changes scattered around the chromosome (Fig. 2): rpoS-ΔG214, trkH-L80Q and lapB-V43G in ENZ1901; kdpD-D460V and rseP-A98V in ENZ1902; and arcA-F172Y, rsxC-I525F, crfC-E693Stop and e14− /icd + in ENZ1903. In the latter strain, excision of the lambdoid prophage e14 restored the icd + sequence that differed from the reference sequence, icdA, by 12 single-nucleotide polymorphisms [18, 19] (Fig. 3).
Fig. 3 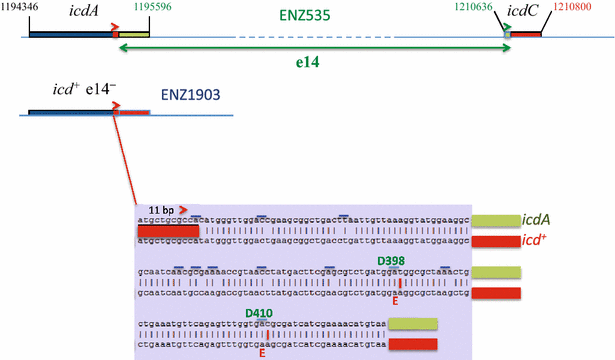
Genetic structures of icd alleles. The icd sequence in ENZ1903 (e14− /icd +) contained 12 single-nucleotide polymorphisms in comparison with the reference sequence, icdA, in ENZ535 (icd::e14). This implicates that a site-specific recombination occurred in the 11-bp direct repeats present in icdA and icdC (≫), which led to the deletion/excision of the prophage e14. Blue bars indicate codons harboring substitutions

Considering the phenotypic and genetic diversity that occurred in evolved populations, a number of questions arise regarding (1) the minimum number of mutations required by a variant to sweep a population [20–23], (2) the types of mutations (loss- or gain-of-function; global regulatory or specific changes) [20–27], and (3) the diversity or convergence of the metabolic changes occurring in evolved strains [28, 29].
In ENZ1901 (phnE + rpoS214 trkH80 lapB43), the trkH80 and rpoS214 mutations similarly affect the metabolic flux
To analyze the effects of the trkH80 and lapB43 mutations present in the evolved strain ENZ1901 (phnE + rpoS214 trkH80 lapB43), we exchanged the rpoS214 allele for the rpoS + allele from the ancestral strain ENZ535, which gave rise to the strain ENZ2043 (phnE + trkH80 lapB43). This strain exhibited a Glg− phenotype, whereas strains that harbored a single phnE + allele (e.g. ENZ1904) exhibited a Glg+ phenotype. Thus, besides the rpoS214 mutation, which decreased the cellular levels of glycogen and of catalase, the trkH80 and/or lapB43 mutation(s) somehow decreased the cellular levels of glycogen.
LapB is an inner-membrane protein that plays a key role in the biogenesis of the glycolipid lipopolysaccharide leaflet in the outer membrane [30, 31]; at first glance, it seemed unlikely that a lapB mutation might affect the glycogen content, a storage compound. In contrast, changes in the activity of TrkH, the primary potassium (K+) transporter, might possibly change the levels of glycogen because the deregulation of K+ homeostasis, which normally occurs in response to an osmotic stress, affects many metabolic processes [32].
To determine whether mutations in trkH could change the Glg phenotype, we transduced the ΔtrkH::kan deletion and the trkH80 evolved mutation (co-transduced with ΔyihL::kan) into the ancestral strain ENZ535. Compared to the parental strain, the ΔtrkH (ENZ2317) and trkH80 (ENZ2163) mutant strains produced more and less glycogen, respectively. Considering that these opposite effects on glycogen contents might reflect changes in K+ transport, and that the rate of transport of K+ is decreased in ΔtrkH mutants [32], we concluded that the rate of transport of K+ could be increased in trkH80 mutants. The TrkH-L80Q mutant protein might therefore mimic the form of TrkH (or of the TrkAEGH complex) that is normally present following an osmotic stress, when the rate of transport of K+ through TrkH transiently increases [32].
Changes in K+ levels may affect the selectivity of sigma factors [32, 33]. To account for the Glg− phenotype of a trkH80 mutant strain, a simple hypothesis was that excess K+ might partially decrease the activity of RpoS, which would reduce the synthesis of glycogen [25]. To test this hypothesis, we used the poxB176::lacZ.Cmr fusion as a reporter of the RpoS activity, and measured the β-galactosidase activity in 1-day old cultures in P- and N-limiting media [10]. The expression of poxB is strictly controlled by the activity of RpoS, which increases poorly in N-starved cells and strongly in P-starved cells [4, 34]. As shown in Table 2, the poxB176::lacZ fusion was barely expressed in strains carrying rpoS-null mutations: ENZ2046 (ΔrpoS) and ENZ2343 (rpoS214 trkH80 lapB43). In contrast, the poxB176::lacZ fusion was normally induced (moderately in N- and strongly in P-limiting media) in the strains ENZ2493 and ENZ2494 that harbored the trkH + and trkH80 alleles, respectively. Therefore, the trkH80 mutation had by itself no effect on the activity of RpoS in Pi-starved cells.
Increased levels of K+ may inhibit the activity of proteins [35–37]. To determine whether the trkH80 mutation could change the activity of enzymes involved in glucose metabolism, we determined the levels of glucose and of acetic acid in spent media during prolonged incubation of trkH + and trkH80 strains in Pi-limiting medium (Fig. 4). Surprisingly, the strains that harbored the trkH80 mutation, independently of the nature of the phnE and lapB alleles (ENZ2163: trkH80; ENZ2395 and ENZ2405: phnE + trkH80; and ENZ2067: phnE + trkH80 lapB43), exhibited similar viability and metabolic pattern as single rpoS mutants during prolonged incubation in monocultures starved for Pi (Fig. 4a–h) [1]. First, trkH80 mutant strains lost viability between days 1 and 6 of incubation (Fig. 4b), while glucose was completely consumed (Fig. 4f) and low levels of acetic acid were excreted (10 mM acetate at pH 6) (Fig. 4d, h). Second, by day 6 of incubation, cells grew on acetic acid (Ace+ phenotype), which was totally detoxified on day 8 of incubation (≤0.02 mM acetate at pH 6.8) (Fig. 4d, h). In contrast, the phnE + lapB43 mutant strain (ENZ2137) behaved as the ancestral strain (ENZ2315: ENZ535 phnE + and ENZ535) (Fig. 4a–h). Collectively, these results indicate that the trkH80 mutation could change the metabolic flux in a similar manner to an rpoS-null mutation, but without affecting the activity of the RpoS factor.

Similar effects of the trkH80 and rpoS-null mutations on the viability and on the metabolic activity of strains incubated in monoculture. Strains were inoculated 1:500 (time zero) into Pi-limiting medium and further incubated. The numbers of CFU (a, b), the pH (c, d) and the concentrations of glucose (e, f) and of acetate (g, h) were determined between days 1 and 24 of incubation. The curves in panels a, c, e, g for the ancestral strain ENZ535, the evolved strain ENZ1901 and the rpoS strain were previously published [1]
Effects of the trkH80, lapB43 and rpoS214 mutations in mixed cultures
To determine the effects of the mutations present in the evolved strain ENZ1901 during prolonged incubation in mixed culture, strains (Kanr) harboring different combinations of the wild type and evolved alleles were added as a minority in cultures of the ancestral strain (Tcr) starved for Pi, further incubated, and the co-cultures were re-diluted 1:50 into fresh Pi-limiting medium on day 9 of incubation and incubated up to day 35 (Fig. 5).

Effects of the trkH80, rpoS214 and lapB43 mutations on the viability of strains incubated in mixed culture. The strains tested as a minority (Kanr) were grown as monocultures in Pi-limiting medium for 1 day, diluted 105-fold into 50 ml of 1-day-old cultures of the ancestral strain ENZ1797 (ENZ535 Tcr) in Pi-limiting medium, and incubated further for 8 days. On day 9 of incubation, 1 ml of mixed culture was added into 50 ml of fresh Pi-limiting medium and incubated further. At the end of the incubation period, the pH and the concentrations of glucose (Glc) and of acetate (Ace) were determined in spent media (concentrations are indicated in mM; the sign < indicates that the concentration was ≤0.02 mM). The viability of the ancestral strain (Tcr) is indicated with dashed lines of different colours for independent experiments. The viability of the test strains (Kanr) -ENZ2000 (a), ENZ1791 and ENZ2166 (b), ENZ2067 (c), ENZ2136 (d), ENZ2395 (e), ENZ2163 (f), ENZ2137 (g), and ENZ2003 and ENZ1984 (h)- is indicated with solid lines
The efficiency of the GPS phenotype (determined on day 5 of incubation) decreased in the order: ≈5 × 107 CFU/ml for phnE + trkH80 lapB43 rpoS214 mutants (ENZ2000) (Fig. 5a) [1]; ≈2 × 107 CFU/ml for phnE + trkH80 lapB43 mutants (ENZ2067 and ENZ2136) (Fig. 5c, d); ≈2 × 106 CFU/ml for phnE + trkH80 (ENZ2395) (Fig. 5e); ≈7 × 104 CFU/ml for phnE + lapB43 mutants (ENZ2137) (Fig. 5g); ≈ 3 × 104 CFU/ml for single trkH80 mutants (ENZ2163) (Fig. 5f); and ≈104 CFU/ml for single phnE + (ENZ1984 and ENZ2003) (Fig. 5h) and parental strains (ENZ1791 and ENZ2166) (Fig. 5b) [1].
When the co-cultures were re-diluted into fresh medium, Kanr strains were generally overtook by the ancestral strain (Tcr) and both strains lost viability owing to the high levels of acetic acid excreted (up to 30 mM acetic acid at pH 4.8, on day 34 of incubation). This notably occurred in the case of the evolved strain ENZ2000 (phnE + trkH80 lapB43 rpoS214) (Fig. 5a). In sharp contrast, triple mutants phnE + trkH80 lapB43 (ENZ2067 and ENZ2136) overtook the ancestral strain, survived prolonged incubation, and afforded a robust cross-protection to the ancestral strain as a result of the detoxification of acetic acid in the incubation medium (≤0.02 mM glucose and acetate at pH 6.8, on day 35 of incubation) (Fig. 5c, d). The phnE + trkH80 double mutants (ENZ2395) sometimes failed to overtake the ancestral strain, which then triggered the death of the whole population during prolonged incubation (Fig. 5e). The phnE + lapB43 strain (ENZ2137) was overtook by the ancestral strain and died, whereas it rapidly evolved RpoS− variants (Fig. 5g).
Therefore, the lapB43 mutation, which had no effect on the metabolic pattern in monoculture (Fig. 4) and only a weak effect on the GPS phenotype, might improve the GPS phenotype and the long-term viability of phnE + trkH80 lapB43 triple mutants in mixed culture. In contrast, the addition of the rpoS214 mutation into the phnE + trkH80 lapB43 strain slightly improved the GPS phenotype but dramatically decreased the long-term viability, which indicates that the presence of the rpoS214 mutation in the evolved strain ENZ1901 (phnE + trkH80 lapB43 rpoS214) was eventually deleterious.
The kdpD-D460V mutation in ENZ1902 triggers a constitutive expression of the kdpFABC operon
All phenotypic traits of the evolved strain ENZ1902 (phnE + kdpD460 rseP98) matched with those of ENZ1901 rpoS + derivatives (phnE + trkH80 lapB43): Glg−, RpoS+ (Table 2), Ace+ in pure culture (Fig. 6a, c, e, g), and GPS+ in mixed culture (Fig. 7a). Therefore, we focused on the kdpD460 mutation, which might affect K+ homeostasis like the trkH80 mutation. KdpD belongs to the Kdp system composed of the high-affinity K+ transporter (KdpFABC) and of the two-component system KdpD/KdpE, in which KdpD is the sensor that signals low levels of K+ in the medium. Under K+-limiting conditions, KdpD phosphorylates KdpE, which induces the kdpFABC operon [32, 38].

Effects of the kdpD460 and rseP98 mutations on the viability and on the metabolic activity of strains incubated in monoculture. Strains were inoculated 1:500 (time zero) into Pi-limiting medium, further incubated, and the numbers of CFU (a, b) in the cultures, the pH (c, d) and the concentrations of glucose (e, f) and of acetate (g, h) in the spent media were determined

The rseP98 mutation triggers a GPS phenotype but reduces the long-term viability of kdpD460 mutants in mixed culture. The experiments were conducted as described in the legend to Fig. 5. The test strains (Kanr) are ENZ2001 (a), ENZ2402 (b), and ENZ2112 (c)
To determine whether the kdpD460 mutation could change the activity of the KdpD/KdpE two-component system, we used a strain carrying a PkdpF::lacZ fusion (ENZ2337) [14]. Transduction of the kdpD460 allele into the reporter strain ENZ2337 increased by 100-fold the levels of β-galactosidase (Fig. 8); similar values are obtained when the reporter strain grows with limiting K+ concentrations [14]. These data indicate that the KdpD-D460V regulator was constitutively active, thereby mimicking the KdpDPi form normally present when cells are starved for K+.

KdpD activity measured with the levels of expression of the Pkdp::lacZ fusion in strains grown in Pi-limiting medium. The strains harbored the kdpD + [ENZ2337 (circle, square, times symbol); ENZ2363 (plus symbol)] and the kdpD460 alleles [ENZ2360 (triangle, inverted triangle, diamond)]. The symbols represent independent experiments. The values were determined between 5 and 24 h of incubation (circle, square, triangle, inverted triangle) or after 24 h of incubation (times symbol, plus symbol, diamond)
The constitutive expression of the Kdp system might simply account for the new “growth inhibitory” (GI) phenotype exhibited by kdpD460 mutant strains during prolonged incubation in mixed cultures. Upon dilution of 9-day old mixed cultures into fresh medium, the evolved strain ENZ2001 (phnE + kdpD460 rseP98 Kanr) (Figs. 7a, 9a) and the reconstructed strain ENZ2402 (phnE + kdpD460 Kanr) (Figs. 7b, 9b) prevented growth of the ancestral strain ENZ1797 (ENZ535 Tcr). Moreover, the evolved strain ENZ1902 (phnE + kdpD460 rseP98, Kanr or Tcr) apparently starved to death the evolved strain ENZ1901 (phnE + rpoS214 trkH80 lapB43, Tcr or Kanr) when the two evolved strains were added as competing minorities in mixed cultures containing the ancestral strain ENZ535 in majority (Fig. 9c, d). In fact, the GI phenotype exhibited by kdpD460 mutant strains might result from the scavenging of K+ through the constitutive activity of the high-affinity Kdp system, which might starve other strains in the population.

The kdpD460 mutant strain prevents the growth of competing strains during prolonged incubation in mixed culture. The strains tested as a minority were grown as monocultures in Pi-limiting medium for 1 day, diluted 105-fold into 50 ml of 1-day-old cultures of the ancestral strain in Pi-limiting medium, and incubated further for 8 days. On day 9 of incubation, 1 ml of mixed culture was added into 50 ml of fresh Pi-limiting medium and incubated further for up to 17 days. The ancestral strain was ENZ1797 (ENZ535 Tcr) when one evolved strain was added as a minority (a, b), and ENZ535 when two evolved strains were added as minorities (c, d)
The rseP-A98V mutation triggers a GPS phenotype
Compared to the evolved strain ENZ1902 (phnE + kdpD460 rseP98), the reconstructed strain ENZ2402 (phnE + kdpD460), which lacked the rseP98 mutation, generally re-consumed acetic acid more efficiently in monoculture (Fig. 6g, h), and in mixed culture (Fig. 7a, b). These results suggest that in the evolved strain ENZ1902, the kdpD460 mutation played a beneficial role, whereas the rseP98 mutation could play a detrimental role in metabolism.
To further characterize the role of the rseP98 mutation in metabolism, we exchanged the kdpD460 mutant allele for the kdpD + allele in ENZ1902, and tested the new strain, ENZ2112 (phnE + rseP98), in monoculture and in mixed culture. In monoculture, the strain ENZ2112 excreted somewhat less acetic acid and consumed less glucose than a phnE + strain (Fig. 6f, h), which confirmed that the rseP98 mutation might cause a slight defect in metabolism during prolonged incubation. Surprisingly, the strain ENZ2112 (phnE + rseP98) exhibited a strong GPS phenotype but did not survive prolonged incubation in mixed culture (Fig. 7c).
RseP is an inner-membrane protein. Its primary role is to contribute to the cleavage of the RseA anti-σE factor, which triggers the stress envelope σE (RpoE) regulon [39, 40]. To determine whether the rseP98 mutation could affect the expression of the RpoE regulon in Pi-starved cells, we transduced the rseP98 and rseP + alleles into a reporter strain carrying the rpoHP3::lacZ fusion, which measured the activity of the σE-RNA polymerase holoenzyme [7]. In an rseP + derivative, the RpoE activity increased significantly at the approach of the stationary phase, especially under Pi starvation conditions (Fig. 10) [2, 7, 41]. A ΔrseA mutation increased by 3.8-fold the levels of expression of the rpoHP3::lacZ fusion in exponential (t = 7 h, OD600 = 0.25) and in stationary phases (t = 24 h) (Table 3), which indicates that the spontaneous induction of the RpoE regulon in Pi-starved cells was independent of the cleavage of RseA [7, 41]. A rseP98 mutation had no effect on the levels of expression of the rpoHP3::lacZ fusion, neither in exponential growth phase nor in stationary phase (Table 3), which rules out a role of the RseP-A98V protein in the expression of the stress envelope σE (RpoE) regulon.

The RpoE activity measured with the levels of expression of the rpoHP3::lacZ fusion is higher in P- than in N-starved cells. The strain ENZ2489 (phnE + rseP +) was inoculated 1:500 (time zero) into P- and N-limiting media and further incubated. The OD600 (dashed lines) and the β-galactosidase activity were determined (solid lines). The symbols represent two independent experiments
Synergy of arcA and rsxC mutations on acetic acid detoxification in ENZ1903
Except for its Glg+ phenotype, the evolved strain ENZ1903 (phnE + arcA-F172Y rsxC-I525F e14− /icd + crfC-E693Stop) exhibited the characteristic phenotypes of kdpD460 and trkH80 mutant strains: Ace+ in monoculture (Fig. 11g), GPS+ (days 1–5) and long-term viability in mixed culture (Fig. 12a).

Synergistic effects of the arcA and rsxC mutations on the viability and metabolic activity of strains incubated in monoculture. The experiments were conducted as described in the legend to Fig. 4

The arcA and rsxC mutations are not sufficient for a long-term viability in mixed culture. The experiments were conducted as described in the legend to Fig. 5. The test strains (Kanr) are ENZ1982 (a) and ENZ2439 (b)
Mutations in arcA, rsxC and icd genes were not unexpected because they could affect the activity of the tricarboxylic acid cycle in different ways:
-
1.
The dual regulator ArcA primarily inhibits the expression of genes implicated in the activity of the tricarboxylic acid cycle (e.g. acnB and fumA) and of the aerobic respiratory chain when the concentration of dissolved oxygen decreases. ArcA also increases the synthesis of CydAB, an enzyme of the aerobic respiratory chain that scavenges low levels of oxygen, which increases the energetic metabolism and extents the protection of cytoplasmic proteins (e.g. FNR) against oxygen at low pO2 [42, 43]. Inactivation of arcA increases the metabolic flux into the tricarboxylic acid cycle in exponentially growing cells [44].
-
2.
RsxC belongs to the Rsx NADPH-dependent reducing system that inactivates SoxR and thus shunts off the SoxRS response to oxidative stress [45]. The SoxRS response comprises acnA and fumC, which encode proteins of the tricarboxylic acid cycle that are resistant to oxidative stress [46]. In exponentially growing cells, a ΔrsxC mutation increases the basal level of expression of the SoxRS regulon primarily because the [2Fe-2S] clusters in SoxR that are spontaneously oxidized cannot be re-reduced [45]. In addition, in arcA mutants, the oxygen-sensitive transcription factor FNR should be inactive, which might increase the cellular levels of SoxR and SoxS [42, 47].
-
3.
During growth on acetate, the activity of Icd (isocitrate dehydrogenase) is decreased as a result of its phosphorylation, which helps to redirect part of the metabolic flux from the tricarboxylic acid cycle towards the glyoxylate shunt [11]. In E. coli K-12, the phage e14 is normally integrated in icd (Fig. 3). Although the sequences of icd in lysogenic (e14+; icdA) and in non-lysogenic strains (e14−; icd +) differ by 12 single-nucleotide polymorphisms, 10 synonymous and two non-synonymous, both forms of the Icd protein exhibit the same activity in vitro. Recent data suggest that excision of the prophage e14 and/or restoration of the icd + gene could somehow decrease or increase the resistance of E. coli K-12 to different stresses [18, 19].
We found evidence that the arcA-F172Y allele encoded an inactive protein. First, we transduced a cydA::lacZ fusion, which is induced by ArcA [13], into the evolved strain ENZ1903 and into the ancestral strain ENZ535. After 24 h of incubation under aerobic, Pi starvation conditions, the levels of β-galactosidase were 3.4-fold lower in ENZ1903 (1787 ± 303 units, n = 2) than in ENZ535 derivatives (6062 ± 401 units, n = 2), which suggests that the evolved strain ENZ1903 might exhibit little, if any, ArcA activity [13]. Second, we exchanged the arcA172 allele in ENZ1903 for a ΔarcA::kan deletion (ENZ2415), which did not change significantly the metabolic patterns of the strains in monocultures (Fig. 11a, c, e, g). Similarly, exchange of the rsxC525 allele in ENZ1903 for a ΔrsxC::kan deletion (ENZ2416) had no significant effect on the metabolic patterns, which suggests that the rsxC-I525F mutation was null or neutral (Fig. 11a, c, e, g).
We transduced the ΔarcA::kan mutation (the kanamycin resistance marker could be eventually excised) and/or the ΔrsxC::kan mutation into the ancestral strain ENZ535, and determined the metabolic patterns of the different strains during incubation in monocultures (Fig. 11). Whereas the ΔrsxC mutation alone (ENZ2414) had no effect on the cellular viability and on the kinetics of production of acetic acid, the ΔarcA mutation alone (ENZ2413) had moderate effects: values were in-between the values obtained for the ancestral strain and the evolved strain ENZ1903 (Fig. 11b, d, f, h). Surprisingly, transduction of the ΔrsxC::kan mutation into the ΔarcA mutant strain (ENZ2434: ENZ535 ΔarcA ΔrsxC) dramatically improved the viability of the strain and its ability to consume the low levels of acetic acid that were previously excreted (Fig. 11b, d, f, h).
A ΔrsxC mutation increases the SoxRS oxidative stress response in ΔarcA mutants
Could the synergy of the ΔarcA and ΔrsxC mutations be explained by an effect of the rsxC mutation on the SoxRS oxidative stress response? To answer this question, we transduced the ΔarcA and/or ΔrsxC mutations into the strain ENZ1843, which carries a soxS::lacZ fusion that is induced by SoxR in its active (oxidized) form [15], and we measured the β-galactosidase activity in exponentially growing cells (6 h of incubation; OD600 = 0.1) and in non-growing cells (24 h of incubation in P-limiting medium).
In exponentially growing cells, the arcA and rsxC mutations exhibited a synergistic effect on the levels of expression of soxS::lacZ, which increased barely in ΔarcA mutants, moderately in ΔrsxC mutants [45], and strongly in ΔarcA ΔrsxC double mutants (3.6-fold compared to wild type cells) (Fig. 13). Surprisingly, in Pi-starved cells, the levels of expression of soxS::lacZ changed in a complex manner (Fig. 13). In the parental strain ENZ1843 (WT), the levels of expression of soxS::lacZ were 2.8-fold higher in non-growing than in growing cells. We have previously shown that Pi-starved cells accumulate oxidative damage in macromolecules as a result of the endogenous production of H2O2 and HO· radicals [4], which might oxidize guanine residues and consequently SoxR bound to the soxS promoter [48]. The levels of expression of soxS::lacZ further increased weakly in ΔarcA mutants, whereas they decreased in ΔrsxC mutants. The latter unexpected result might reflect the fact that in the absence of the RsxC activity, oxidized [Fe-S] clusters in SoxR may eventually lose Fe [45], which generates inactive apo-SoxR forms. Despite this caveat, the levels of expression of soxS::lacZ increased by 1.5-fold in ΔarcA ΔrsxC double mutants compared to ΔarcA single mutants (Fig. 13), which indicates that the ΔrsxC mutation could afford a protection to ΔarcA mutants starved for Pi through a higher level of expression of the SoxRS oxidative stress response.

The combination of the arcA and rsxC mutations increases the levels of expression of the soxS::lacZ fusion. Strain ENZ1843 (soxS::lacZ) and derivatives ENZ2420 (ΔarcA), ENZ2421 (ΔrsxC) and ENZ2436 (ΔarcA ΔrsxC) were inoculated (0.1 ml) into 50 ml of Pi-limiting medium and β-galactosidase activities were determined in exponentially growing cells (6 h of incubation; OD600 = 0.1; n = 2) (ex) and in non-growing cells (24 h of incubation; n = 3) (P-). The values are the means ± standard deviations for n independent experiments
The arcA and rsxC mutations are not sufficient for a long-term viability in mixed culture
The constructed strain ENZ2439 (ENZ535 phnE + ΔarcA ΔrsxC::kan), added in minority in a mixed culture, grew vigorously but died more rapidly than the evolved strain ENZ1982 (ENZ1903 ΔlacY::kan) between days 6 and 9 of incubation (Fig. 12a, b). Moreover, following the dilution of the mixed cultures into fresh medium on day 9 of incubation, the reconstructed strain ENZ2439 grew slowly and died off (Fig. 12b), whereas the evolved strain ENZ1982 overtook the population and survived prolonged incubation (Fig. 12a). A simple interpretation of these results is that the icd + and/or crfC693 alleles, present in the evolved strain (phnE + arcA172 rsxC525 e14− /icd + crfC693), but not in the reconstructed strain ENZ2439 (phnE + ΔarcA ΔrsxC icd::e14), might help cells to tolerate a stress that built up in mixed culture.
Discussion
It is generally thought that cells that evolve under nutrient limitation conditions primarily accumulate mutations that decrease the activity of the RpoS (σs) factor [18, 20–22, 25, 26, 28, 29, 49]. We show here that in strains that evolved in serial batch cultures under aerobic, Pi starvation conditions, rpoS-null mutations, which were first beneficial, were eventually detrimental. In contrast, we identified novel combinations of mutations that were beneficial to the long-term viability of the cells.
K+ homeostasis and metabolic flux
The trkH80 and kdpD460 mutations played a key role in the survival of the evolved strains ENZ1901 (phnE + rpoS-ΔG214 trkH-L80Q lapB-V43G) and ENZ1902 (phnE + kdpD-D460V rseP-A98V), respectively. The trkH80 and kdpD460 mutations triggered a constitutive expression of the low-affinity high-rate Trk system and of the high-affinity low-rate Kdp system, respectively.
The trkH-L80Q mutation was previously identified in strains that evolved in Pi-limited chemostat cultures and in serial batch cultures containing the antibiotic gentamicin sulfate [25, 50]. In the strain evolved in Pi-limited chemostat, the trkH-L80Q mutation was not further characterized because of the presence of numerous mutations [25]. In strains resistant to gentamicin, it was suggested that excess K+ could prevent the transport of the antibiotic inside the cells [50, 51]. The kdpD-D460V constitutive mutation was not previously described but the KdpD-D460V evolved protein is reminiscent of the KdpD-D474A engineered protein, which cannot sense K+ in the periplasm and thus constitutively activates the KdpE regulator of the kdpFABC operon [38].
How excess K+ could change the metabolism of glucose and of acetic acid in evolved strains? Potassium glutamate constitutes the most abundant ions inside the cell; K+ transport is linked to Pi transport and to internal pH (K+ transport inside the cell triggers a net H+ excretion) [32, 33, 36, 38, 52]. The activity of the K+ transporters can rapidly increase when the osmotic pressure of the medium increases and when the concentration of K+ in the medium decreases [32, 36], but excess K+ may be eventually toxic [32]. Interestingly, IlvBN, the first enzyme in the pathway of biosynthesis of branched-chain amino acids is especially sensitive to allosteric inhibition by K+ [37]. Because ilvB evolved through duplication/deletion events from poxB [53], it is tempting to speculate that both IlvB and PoxB might be inhibited by excess K+, which would direct the metabolic flux from pyruvate towards the tricarboxylic acid cycle rather than towards the synthesis of branched-chain amino acids and especially acetic acid in Pi-starved cells (Fig. 1).
Changes in outer membrane protein activities
We found evidence that the lapB43 mutation, present in the evolved strain ENZ1901, might help growth on glucose and organophosphates (GPS phenotype), and eventually on acetate and Pi (Ace+ phenotype). LapB is an inner membrane protein that negatively controls the biosynthesis of the lipopolysaccharide [30, 31]. Because the lipopolysaccharide is negatively charged as a result of the presence of two phosphates in lipid A [54], we suggest that the lipopolysaccharide might hamper the binding to the outer membrane of negatively charged products such as Pi and organophosphates. Therefore, we speculate that lapB43 might be a gain-of-function mutation that would decrease the levels of the lipopolysaccharide, which might increase the binding and the diffusion of Pi and organophosphates required for the expression of the GPS phenotype (Fig. 14). The binding of P compounds to the outer membrane may be critical in rpoS + cells in which the induction of the Pho regulon and the synthesis of P-compound scavengers are very transient [25]. In this light, the acquisition of the rpoS214-null mutation in the phnE + lapB43 strain might favor the GPS phenotype in two different ways: by rerouting the metabolic flux towards the tricarboxylic acid cycle [1] and by extending the expression of the Pho regulon [25]. Unfortunately, the rpoS214 mutation reduced the long-term viability of the evolved strain ENZ1901 in mixed culture; this may reflect a defect in acetic acid resistance through the so-called RpoS-dependent amino acid-independent “acid resistance system 1” [55].

Schematic overview of possible changes in ENZ1901 and ENZ1902. BCAA branched-chain amino acids, LPS lipopolysaccharide, PDH pyruvate dehydrogenase, PST phosphate transport, PYR pyruvate, R-O-P organophosphate, TCA tricarboxylic acid
The rseP98 mutation, present in ENZ1902, triggered growth on glucose and organophosphates but hampered growth on acetic acid and Pi, which threatened the long-term viability of the cells in mixed culture. Although the primary role of RseP is to cleave the anti-sigma E factor, RseA [39], the rseP98 mutation had no effect on the level of expression of the RpoE (σE) regulon, which was normally induced in Pi-starved cells [5, 7]. Because inner membrane metalloproteins such as FtsH and RseP may exhibit dual functions in membrane protein biogenesis, as protease and as chaperone [56–58], it is tempting to speculate that the RseP-A98V protein might affect the folding of outer membrane proteins, which might increase the PhnE-dependent scavenging of organophosphates and decrease the PhoE-dependent scavenging of Pi (Fig. 14). Such trade-off would be reminiscent of the apparent changes in activities of the porins OmpC and PhoE, which occur in chemostat cultures limited in glucose [28].
Resistance to oxidative and acetic acid stresses
The arcA172 mutation might play a key role in the metabolic changes that occurred in the evolved strain ENZ1903. The global regulator ArcA progressively inhibits the transcription of genes of the tricarboxylic acid cycle when the concentration of dissolved oxygen decreases, which redirects the metabolic flux towards the production of weak acids such as acetic acid [44]: the inactivation of ArcA might thus maintain the activity of the tricarboxylic acid cycle in Pi-starved cells, which might decrease the production of acetic acid and allow its re-consumption (Fig. 1). Although different arcA mutations occurred in strains that evolved under nutrient stress conditions, the arcA172 mutation, which affects the DNA-binding domain of ArcA, was not previously described [21, 22, 26].
A single ΔarcA mutation moderately decreased the production of acetic acid under aerobic, Pi starvation conditions, which suggests that the tricarboxylic acid cycle was poorly expressed. It is generally thought that the limited activity of the tricarboxylic acid cycle in arcA mutants might result from an insufficient activity of the aerobic respiratory chain, which would limit the re-oxidation of NADH; excess NADH may inhibit the activity of the citrate synthase, GltA, which controls the entry into the tricarboxylic acid cycle [11, 59, 60] (Fig. 1). As our results revealed novel synergistic effects between the ΔarcA and ΔrsxC mutations on the metabolism of acetate and on the expression of the SoxRS oxidative stress response, we would like to suggest that metabolic defects in ΔarcA mutants would result not from the inhibition of enzyme activities by excess NADH but rather from the degradation of enzymes of the tricarboxylic acid cycle (e.g. AcnB and FumA) by reactive oxygen species adventitiously produced by NADH dehydrogenases [4] (Fig. 1).
In mixed culture, the evolved strain ENZ1903 (phnE + arcA172 rsxC525 e14−/icd + crfC693) survived better than the ENZ535 phnE +ΔarcA ΔrsxC reconstructed strain, which suggests that the e14−/icd + and/or crfC693 genetic changes might protect the strain ENZ1903. Interestingly, strains evolving at pH 4.8 accumulate mutations in icd, including an icd-D398E mutation [61]. We realized that this mutation was identical to a single-nucleotide polymorphism induced by the excision of the prophage e14 from icd: the icdA (icd::e14) and icd + (e14−) alleles differed by 12 single-nucleotide polymorphisms, 10 synonymous and 2 non-synonymous including the D398E change (Fig. 3). Although the enzymes encoded by the icdA and icd + alleles exhibit the same activity in vitro, it is likely that active enzymes were produced at different levels in vivo as a result of changes in codon usage, which may affect the rate of synthesis and the folding of the proteins [62, 63]. We propose that the isocitrate dehydrogenase activity might increase as a result of the e14− /icd + genetic changes, which might increase the levels of α-ketoglutarate, of glutamate (synthesized from α-ketoglutarate), and of NADH (produced by the NAD-dependent α-ketoglutarate dehydrogenase): glutamate may be used by the RpoS-dependent GadB decarboxylase [10], which may consume protons inside the cell, and the re-oxidation of NADH by the aerobic respiratory chain may trigger the excretion of protons (Fig. 1). Both processes might help the evolved strain ENZ1903 to resist internal acidification by acetic acid (pH 4.8) that was excreted into the medium by the ancestral strain present in majority in mixed culture between days 1 and 9 of incubation. In this light, it is tempting to speculate about how the truncation of CrfC (crfC-E693Stop), which normally tethers clamp-replication forks complexes [64], could favor the e14−/icd + genetic changes: the CrfC693 mutant protein might destabilize the replication forks [64], generate excess single-stranded DNA, activate the RecA protein, and trigger the excision of the lambdoid prophage e14 (e14−) [65, 66].
Parallel evolutions that change acetate producers in acetate consumers
Collectively, our results revealed that to survive during prolonged incubation under aerobic, Pi starvation conditions, evolved strains acquired several types of mutations that helped them to grow on glucose and organophosphates (GPS phenotype), to produce low levels of acetic acid, and to combat endogenous oxidative stress and exogenous acid stress. Then, evolved strains that overtook the population could grow on acetic acid and Pi previously excreted (Ace+ phenotype), thereby detoxifying the medium [5].
Based on the dynamics of evolving populations (generally, RpoS− strains spread first, followed by Glg− and eventually Glg+ strains) [1], we would like to suggest a possible scenario for the time course of spreading of evolving strains. It is likely that the phnE + change was primarily selected because the 8-bp deletion in phnE, which results from a slippage between direct repeats during replication, occurs with a high frequency [1]. Then, acquisition of the lapB43 and especially rpoS214 mutations, which triggered growth and thus DNA replication under Pi starvation conditions, might increase the probability of acquiring the trkH80 constitutive mutation, thereby generating the strain ENZ1901 that exhibited a high metabolic rate [1]. A similar evolutionary process might occur with the acquisition of the phnE + and rseP98 mutations, which triggered a GPS phenotype, and ultimately of the kdpD460 constitutive mutation, thereby generating the strain ENZ1902. Although the evolved strains ENZ1901 (RpoS−) and ENZ1902 (Glg−) were eventually doomed to die, these strains could provide a transient protection to the population by consuming glucose while producing low levels of acetic acid, which could provide sufficient time for the progressive accumulation of the arcA172 and rsxC525 mutations, and then the crfC93 and e14− /icd + genetic changes, thereby generating the strain ENZ1903. Eventually, during prolonged incubation in detoxified spent medium at pH 7, phnE + single mutant strains (ENZ1904 and ENZ1905) could sweep the population, thereby restoring the ancestral metabolism.
Useful strains
Considering the characteristics of the evolved (ENZ1903) and reconstructed strains (trkH80 lapB43 and kdpD460), we suggest that these strains could be useful in human health and in industrial production of bacterial products.
As previously suggested, E. coli strains that evolved under aerobic, Pi starvation conditions might help to colonize the human intestinal tract, in which both Pi and glutamate are reabsorbed [1]. In this light, it would be interesting to determine whether evolved strains could exhibit probiotic effects, alone or in consortium [67].
The consumption of energy and amino acids for growth and the generation of metabolic stresses (e.g. envelope stress) can limit the production and excretion of proteins and chemicals. To resolve these problems, it has been suggested to stop growth but not protein synthesis, and to use engineered strains (e.g. pta ackA poxB and arcA iclR mutants) that produce less acetic acid and are more tolerant to stress [68–71]. We would like to suggest that the evolved strain ENZ1903 (phnE + arcA172 rsxC525 crfC693 e14− /icd +) incubated in batch culture under aerobic, Pi starvation conditions could afford a platform for producing toxic proteins and chemicals because of two major characteristics: (1) the sustained activity of the tricarboxylic acid cycle in stationary phase, which helps to produce energy and building blocks rather than acetic acid, and (2) the spontaneous induction of several stress responses, i.e. the SoxRS oxidative stress response and a novel e14− /icd + acid resistance system, which are specifically induced in ENZ1903, in addition to the Cpx and RpoE envelope stress responses and the RpoS general stress response, which are normally induced in Pi-starved cells [2].
Methods
Strains
The E. coli K-12 laboratory strain MG1655 is very close to commensal strains [72]. Among several MG1655 isolates [17], we chose the strain CF1648, therein called ENZ535 (Table 1), because it exhibited a high growth rate and a high survival rate during prolonged incubation in rich medium, in glycerol-minimal medium under aerobic conditions, and in glucose-minimal medium under aerobic and anaerobic conditions [1, 10, 73]. Strains of the Keio collection [16] were used as donors to transduce single-gene in-frame deletions replaced with a kanamycin-resistance cassette (Kanr). Transductions were performed with phage P1vir and cells grown in LB5 medium containing 5 g/l NaCl [73, 74]. To construct Kans derivatives of deletion mutants, the kan cassette flanked by the FLP-recombination-target sites was removed by introducing the FLP recombinase-expressing plasmid pCP20 at 32 °C and purifying clones at 42 °C [16]. We co-transduced evolved mutations with Kanr markers from the Keio collection: trkH with ΔyihL::kan, kdpD with ΔybfH::kan, and rseP with ΔyaeH::kan. The ΔlacIZ mutants were constructed as previously described [10]. Spontaneous PhnE+ strains were selected on MOPS minimal medium plates containing 0.05 mM methyl phosphonate as the sole source of P [1].
Before we sequenced the genomes of the evolved strains, we exchanged rpoS alleles for rpoS + in ENZ1901 and ENZ1902. Practically, we transduced the cysC::Tn10 tetracycline-resistance marker (Tcr; 7 kb apart from rpoS, frequency of co-transduction of 35 %) from ENZ2005 (ENZ535 cysC::Tn10 ΔrpoS::kan) into evolved strains, selected cysC::Tn10 ΔrpoS::kan (Tcr Kanr) strains, and transduced the cysC + rpoS + region from the ancestral strain ENZ535 (Cys+ Kans). The evolved strains ENZ1901 (RpoS−) gave rise to ENZ2043 (Glg−), and the evolved strains ENZ1902 (Glg−) to ENZ2044 (Glg−). In the latter case, the strains ENZ1902 and ENZ2044 were indeed identical (rpoS +) as revealed by sequencing of the genomes.
Media and culture conditions
The MOPS minimal medium used for the pre-cultures contained notably 40 mM MOPS (pH 7.4), 86 mM NaCl, 9.5 mM NH4Cl, 20 mM glucose, and 5 mM K2HPO4. The P-limiting MOPS minimal medium contained 40 mM MOPS (pH 7.4), 86 mM NaCl, 9.8 mM KCl, 9.5 mM NH4Cl, 40 mM glucose, and 0.1 mM K2HPO4. The N-limiting MOPS minimal medium contained 40 mM MOPS (pH 7.4), 86 mM NaCl, 2 mM NH4Cl, 40 mM glucose, and 5 mM K2HPO4 [10].
To perform monocultures, strains grown for 24 h in MOPS medium were inoculated (0.1 ml) into 50 ml of P- or N-limiting medium in 500-ml Erlenmeyer flasks (time zero) and incubated with agitation at 150 rpm in a covered water bath rotary shaker at 37 °C. To perform mixed cultures, the strains tested as a minority were grown as monocultures in P-limiting medium for 1 day, diluted 103-fold in MOPS0 buffer (MOPS medium without P, N and C source), added (0.5 ml) into 50 ml of 1-day-old cultures of the ancestral strain in P-limiting medium, and incubated for 8 days. On day 9 of incubation, 1 ml of mixed culture was added into 50 ml of fresh P-limiting medium and incubated further for up to 30 days. In order to distinguish the different strains in mixed cultures, we transduced the lacY::Tn10 (Tcr) and ΔlacY::kan (Kanr) mutations [1].
Determination of RpoS, Glg and PhnE phenotypes
The RpoS− (Glg− KatE−) and Glg− (Glg− KatE+) phenotypes were determined on isolated colonies grown on LB10 medium plates containing 10 g/l NaCl as previously described [1, 74]. The PhnE+ phenotype was determined on isolated colonies grown on glucose MOPS minimal medium plates containing 0.05 mM methyl phosphonate as the sole source of P [1].
Measurement of cell viability
To assess cell viability, serial dilutions were prepared in M9 buffer and aliquots (10 μl) were spotted in triplicate onto LB10 medium plates, which were spread with 2000 units catalase and might contain 30 μg/ml kanamycin or 12 μg/ml tetracycline [1, 74]. In the figures, the values of 10 CFU/ml in parentheses indicate that no CFU were detected when 5 × 20-μl portions of the cultures were directly plated.
Levels of glucose and of acetic acid
The pH of the culture supernatants were determined at 25 °C, adjusted to pH 7 and the concentrations of glucose and acetate were determined by enzymatic tests (R-Biopharm) [1].
Measurement of β-galactosidase activity
The β-galactosidase activity was determined as previously described [73, 74]. Units of β-galactosidase were expressed per OD600 [74].
DNA sequencing
Whole-genome sequencing was performed with genomic DNA isolated with the “Wizard Genomic DNA Purification Kit” (Promega). Genomic DNA library preparation (Insert size: ~330 bp; Tiles: 120), sequencing (“Kit v5”, Illumina) with the “Genome Analyzer GA-IIx Illumina” (74 cycles), and data analysis (CASAVA-1.8.2) were performed by Imagif (CNRS). Tablet (1.12.13.26) was used for the visualization of sequence assemblies [75].
Sequencing of the genes of interest in the evolved and constructed strains was performed from PCR fragments generated with the “PCR Master Mix” (Promega) from colonies. PCR products were purified using a “QIAquick” PCR purification kit (Qiagen) and sequencing was performed by Beckman Coulter Genomics.
References
Guillemet ML, Moreau PL. Activation of the cryptic PhnE permease promotes rapid adaptive evolution in a population of Escherichia coli K-12 starved for phosphate. J Bacteriol. 2012;194:253–60.
Moreau PL. Protective role of the RpoE (σE) and Cpx envelope stress responses against gentamicin killing on nongrowing Escherichia coli incubated under aerobic, phosphate starvation conditions. FEMS Microbiol Lett. 2014;357:151–6.
Hsieh Y-J, Wanner BL. Global regulation by the seven-component Pi signaling system. Curr Opin Microbiol. 2013;13:198–203.
Moreau PL. Diversion of the metabolic flux from pyruvate dehydrogenase to pyruvate oxidase decreases oxidative stress during glucose metabolism in nongrowing Escherichia coli cells incubated under aerobic, phosphate starvation conditions. J Bacteriol. 2004;186:7364–8.
Guillemet ML, Moreau PL. Fur-dependent detoxification of organic acids by rpoS mutants during prolonged incubation under aerobic, phosphate starvation conditions. J Bacteriol. 2008;190:5567–75.
Battesti A, Majdalani N, Gottesman S. The RpoS-mediated general stress response in Escherichia coli. Ann Rev Microbiol. 2011;65:189–213.
Gopalkrishnan S, Nicoloff H, Ades SE. Co-ordinated regulation of the extracytoplasmic stress factor, sigma E, with other Escherichia coli sigma factors by (p)ppGpp and DksA may be achieved by specific regulation of individual holoenzymes. Mol Microbiol. 2014;93:479–93.
Weber H, Polen T, Heuveling J, Wendisch VF, Hengge R. Genome-wide analysis of the general stress response network in Escherichia coli: σS-dependent genes, promoters, and sigma factor selectivity. J Bacteriol. 2005;187:1591–603.
Korshunov S, Imlay JA. Detection and quantification of superoxide formed within the periplasm of Escherichia coli. J Bacteriol. 2006;188:6326–34.
Moreau PL. The lysine decarboxylase CadA protects Escherichia coli starved of phosphate against fermentation acids. J Bacteriol. 2007;189:2249–61.
Cronan JE, Laporte D. Tricarboxylic acid cycle and glyoxylate bypass. EcoSal Plus. 2013. doi:10.1128/ecosalplus.3.5.2.
Wolfe AJ. The acetate switch. Microb Mol Biol Rev. 2005;69:12–50.
Bekker M, Alexeeva S, Laan W, Sawers G, Teixeira de Mattos J, Hellingwerf K. The ArcBA two-component system of Escherichia coli is regulated by the redox state of both the ubiquinone and the menaquinone pool. J Bacteriol. 2010;192:746–54.
Fried L, Lassak J, Jung K. A comprehensive toolbox for the rapid construction of lacZ fusion reporters. J Microbiol Methods. 2012;91:537–43.
Nunoshiba T, Hidalgo E, Li Z, Demple B. Negative autoregulation by the Escherichia coli SoxS protein: a dampening mechanism for the soxRS redox stress response. J Bacteriol. 1993;175:7492–4.
Baba T, Ara T, Hasegawa M, Takai Y, Okumura Y, Baba M, Datsenko KA, et al. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol Syst Biol. 2006;2006(2):0008.
Freddolino PL, Amini S, Tavazoie S. Newly identified genetic variations in common Escherichia coli MG1655 stock cultures. J Bacteriol. 2012;194:303–6.
Charusanti P, Conrad TM, Knight EM, Venkataraman K, Fong NL, Xie B, Gao Y, Palsson BØ. Genetic basis of growth adaptation of Escherichia coli after deletion of pgi, a major metabolic gene. PLoS Genet. 2010;6(11):e1001186.
Wang X, Kim Y, Ma Q, Hong SH, Pokusaeva K, Sturino JM, Wood TK. Cryptic prophages help bacteria cope with adverse environments. Nat Commun. 2010;1:147. doi:10.1038/ncomms1146.
Zinser ER, Schneider D, Blot M, Kolter R. Bacterial evolution through the selective loss of beneficial genes: trade-offs in expression involving two loci. Genetics. 2003;164:1271–7.
Plucain J, Hindré T, Le Gac M, Tenaillon O, Cruveiller S, Médigue C, et al. Epistasis and allele specificity in the emergence of a stable polymorphism in Escherichia coli. Science. 2014;343:1366–9.
Saxer G, Krepps MD, Merkley ED, Ansong C, Deatherage Kaiser BL, Valovska M-T, et al. Mutations in global regulators lead to metabolic selection during adaptation to complex environments. PLoS Genet. 2014;10(12):e1004872.
Lang GI, Rice DP, Hickman MJ, Sodergren E, Weinstock GM, Botstein D, Desai MM. Pervasive genetic hitchhiking and clonal interference in forty evolving yeast populations. Nature. 2013;500:571–4.
Johnson MD, Bell J, Clarke K, Chandler R, Pathak P, Xia Y, et al. Characterization of mutations in the PAS domain of the EvgS sensor kinase selected by laboratory evolution for acid resistance in Escherichia coli. Mol Microbiol. 2014;93:911–27.
Wang L, Spira B, Zhou Z, Feng L, Maharjan RP, Li X, et al. Divergence involving global regulatory gene mutations in an Escherichia coli population evolving under phosphate limitation. Genome Biol Evol. 2010;2:478–87.
Herron MD, Doebeli M. Parallel evolutionary dynamics of adaptive diversification in Escherichia coli. PLoS Biol. 2013;11(2):e1001490.
Hottes AK, Freddolino PL, Khare A, Donnell ZN, Liu JC, Tavazoie S. Bacterial adaptation through loss of function. PLoS Genet. 2013;9(7):e1003617.
Maharjan R, Seeto S, Notley-McRobb L, Ferenci T. Clonal adaptive radiation in a constant environment. Science. 2006;313:514–7.
Tenaillon O, Rodriguez-Verdugo A, Gaut RL, McDonald P, Bennett AF, Long AD, Gaut BS. The molecular diversity of adaptive convergence. Science. 2012;335:457–61.
Klein G, Kobylak N, Lindner B, Stupak A, Raina S. Assembly of lipopolysaccharide in Escherichia coli requires the essential LapB heat shock protein. J Biol Chem. 2014;289:14829–53.
Mahalakshmi S, Sunayana MR, SaiSree L, Reddy M. yciM is an essential gene required for regulation of lipopolysaccharide synthesis in Escherichia coli. Mol Microbiol. 2014;91:145–57.
Altendorf K, Booth IR, Gralla J, Greie J-C, Rosenthal AZ, Wood JM. Osmotic stress. EcoSal Plus. 2013. doi:10.1128/ecosalplus.5.4.5.
Lee C-R, Cho S-H, Kim H-J, Kim M, Peterkofsky A, Seok Y-J. Potassium mediates Escherichia coli enzyme IIANtr-dependent regulation of sigma factor selectivity. Mol Microbiol. 2010;78:1468–83.
Zafar MA, Carabetta VJ, Mandel MJ, Silhavy TJ. Transcriptional occlusion caused by overlapping promoters. Proc Natl Acad Sci USA. 2014;111:1557–61.
Heermann R, Weber A, Mayer B, Ott M, Hauser E, Gabriel G, et al. The universal stress protein UspC scaffolds the KdpD/KdpE signaling cascade of Escherichia coli under salt stress. J Mol Biol. 2008;386:134–48.
Pflüger-Grau K, Görke B. Regulatory roles of the bacterial nitrogen-related phosphotransferase system. Trends Microbiol. 2010;18:205–14.
Reaves ML, Rabinowitz JD. Characteristic phenotypes associated with ptsN-null mutants in Escherichia coli K-12 are absent in strains with functional ilvG. J Bacteriol. 2011;193:4576–81.
Laermann V, Cudic E, Kipschull K, Zimmann P, Altendorf K. The sensor kinase KdpD of Escherichia coli senses external K+. Mol Microbiol. 2013;88:1194–204.
Ades SE. Regulation by destruction: design of the σE envelope stress response. Curr Opin Microbiol. 2008;11:535–40.
Ades SE, Grigorova IL, Gross CA. Regulation of the alternative sigma factor σE during initiation, adaptation, and shutoff of the extracytoplasmic heat shock response in Escherichia coli. J Bacteriol. 2003;185:2512–9.
Costanzo A, Nicoloff H, Barchinger SE, Banta AB, Gourse RL, Ades SE. ppGpp and DksA likely regulate the activity of the extracytoplasmic stress factor σE in Escherichia coli by both direct and indirect mechanisms. Mol Microbiol. 2008;67:619–32.
Rolfe MD, Ocone A, Stapleton MR, Hall S, Trotter EW, Poole RK, et al. Systems analysis of transcription factor activities in environments with stable and dynamic oxygen concentrations. Open Biol. 2012;2:120091.
Park DM, Akhtar MS, Ansari AZ, Landick R, Kiley PJ. The bacterial response regulator ArcA uses a diverse binding site architecture to regulate carbon oxidation globally. PLoS Genet. 2013;9(10):e1003839.
Perrenoud A, Sauer U. Impact of global transcriptional regulation by ArcA, ArcB, Cra, Crp, Cya, Fnr, and Mlc on glucose catabolism in Escherichia coli. J Bacteriol. 2005;187:3171–9.
Koo M-S, Lee J-H, Rah S-Y, W-s Yeo, Lee J-W, Lee K-L, et al. A reducing system of the superoxide sensor SoxR in Escherichia coli. EMBO J. 2003;22:2614–22.
Imlay JA. Cellular defenses against superoxide and hydrogen peroxide. Ann Rev Biochem. 2008;77:755–76.
Kumar R, Shimizu K. Transcriptional regulation of main metabolic pathways of cyoA, cydB, fnr, and fur gene knockout Escherichia coli in C-limited and N-limited aerobic continuous cultures. Microb Cell Fact. 2011;10:3.
Lee PE, Demple B, Barton JK. DNA-mediated redox signaling for transcriptional activation of SoxR. Proc Natl Acad Sci USA. 2009;106:13164–8.
Maharjan R, Zhou Z, Ren Y, Li Y, Gaffé J, Schneider D, et al. Genomic identification of a novel mutation in hfq that provides multiple benefits in evolving glucose-limited populations of Escherichia coli. J Bacteriol. 2010;192:4517–21.
Lázár V, Singh GP, Spohn R, Nagy I, Horvath B, Hrtyan M, et al. Bacterial evolution of antibiotic hypersensitivity. Mol Syst Biol. 2013;9:700.
Taber HW, Mueller JP, Miller PF, Arrow AS. Bacterial uptake of aminoglycoside antibiotics. Microbiol Rev. 1987;51:439–57.
Lüttmann D, Göpel Y, Görke B. The phosphotransferase protein EIIANtr modulates the phosphate starvation response through interaction with histidine kinase PhoR in Escherichia coli. Mol Microbiol. 2012;86:96–110.
Jensen RA. Evolution of metabolic pathways in enteric bacteria. In: Neidhardt FC, Curtiss III R, Ingraham JL, Lin ECC, Low KB, Magasanick B, Resnikoff WS, Riley M, Schaechter M, Umbarger HE, editors. Escherichia coli and Salmonella: cellular and molecular biology. Washington, DC: ASM Press; 1996. p. 2649–62.
Kato A, Chen HD, Latifi T, Groisman EA. Reciprocal control between a bacterium’s regulatory system and the modification status of its lipopolysaccharide. Mol Cell. 2012;47:897–908.
Price SB, Wright JC, DeGraves FJ, Castanie-Cornet MP, Foster JW. Acid resistance systems required for survival of Escherichia coli O157:H7 in the bovine gastrointestinal tract and in apple cider are different. Appl Environ Microbiol. 2004;70:4792–9.
Douchin V, Bohn C, Bouloc P. Down-regulation of porins by a small RNA bypasses the essentiality of the regulated intramembrane proteolysis protease RseP in Escherichia coli. J Biol Chem. 2006;281:12253–9.
Hizukuri Y, Akiyama Y. PDZ domains of RseP are not essential for sequential cleavage of RseA or stress-induced σE activation in vivo. Mol Microbiol. 2012;86:1232–45.
Li W, Rao DK, Kaur P. Dual role of the metalloprotease FtsH in biogenesis of the DrrAB drug transporter. J Biol Chem. 2013;288:11854–64.
Prüss BM, Nelms JM, Park C, Wolfe AJ. Mutations in NADH: ubiquinone oxidoreductase of Escherichia coli affect growth on mixed amino acids. J Bacteriol. 1994;176:2143–50.
Vemuri GN, Altman E, Sangurdekar DP, Khodursky AB, Eiteman MA. Overflow metabolism in Escherichia coli during steady-state growth: transcriptional regulation and effect of the redox ratio. Appl Environ Microbiol. 2006;72:3653–61.
Harden MM, He A, Creamer K, Clark MW, Hamdallah I, Martinez KA, et al. Acid-adapted strains of Escherichia coli K-12 obtained by experimental evolution. Appl Environ Microbiol. 2015;81:1932–41.
Goodman DB, Church GM, Kosuri S. Causes and effects of N-terminal codon bias in bacterial genes. Science. 2013;342:475–9.
Kim SJ, Yoon JS, Shishido H, Yang Z, Rooney LA, Barral JM, Skach WR. Translational tuning optimizes nascent protein folding in cells. Science. 2015;348:444–8.
Ozaki S, Matsuda Y, Keyamura K, Kawakami H, Noguchi Y, Kasho K, et al. A replicase clamp-binding dynamin-like protein promotes colocalization of nascent DNA strands and equipartitioning of chromosomes in Escherichia coli. Cell Rep. 2013;4:985–95.
Moreau PL. Effects of overproduction of single-stranded DNA-binding protein on RecA-dependent processes in Escherichia coli. J Mol Biol. 1987;194:621–34.
Moreau PL, Fanica M, Devoret R. Cleavage of λ repressor and induction of RecA protein synthesis elicited by aflatoxin B1 metabolites in Escherichia coli. Carcinogenesis. 1980;1:837–48.
Bron PA, van Baarlen P, Kleerebezem M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat Rev Microbiol. 2012;10:66–78.
Causey TB, Shanmugam KT, Yomano LP, Ingram LO. Engineering Escherichia coli for efficient conversion of glucose to pyruvate. Proc Natl Acad Sci USA. 2004;101:2235–40.
Waegeman H, Beauprez J, Moens H, Maertens J, DeMey M, Foulquié-Moreno M, et al. Effect of iclR and arcA knockouts on biomass formation and metabolic fluxes in Escherichia coli K12 and its implications on understanding the metabolism of Escherichia coli BL21 (DE3). BMC Microbiol. 2011;11:70.
Foo JL, Jensen HM, Dahl RH, George K, Keasling JD, Lee TS, et al. Improving microbial biogasoline production in Escherichia coli using tolerance engineering. mbio. 2014;5(6):e01932-14.
Mahalik S, Sharma AK, Mukherjee KJ. Genome engineering for improved recombinant protein expression in Escherichia coli. Microb Cell Fact. 2014;13:177.
Vital M, Chai B, Ostman B, Cole J, Konstantinidis KT, Tiedje JM. Gene expression analysis of Escherichia coli strains provides insights into the role of gene regulation in diversification. ISME J. 2015;9:1130–40.
Gérard F, Dri A-M, Moreau PL. Role of Escherichia coli RpoS, LexA and H-NS global regulators in metabolism and survival under aerobic, phosphate-starvation conditions. Microbiology. 1999;145:1547–62.
Miller JH. Experiments in molecular genetics. New York: Cold Spring Harbor Lab; 1972.
Milne I, Bayer M, Cardle L, Shaw P, Stephen G, Wright F, Marshall D. Tablet-next generation sequence assembly visualization. Bioinformatics. 2010;26:401–2.
Authors’ contributions
LL carried out the molecular genetic studies. PLM conceived of the study, performed the experiments and wrote the manuscript. Both authors read and approved the final manuscript.
Acknowledgements
We thank A. Battesti for comments on the manuscript. The Centre National de la Recherche Scientifique and Aix-Marseille Université provided funding.
Competing interests
Both authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Moreau, P.L., Loiseau, L. Characterization of acetic acid-detoxifying Escherichia coli evolved under phosphate starvation conditions. Microb Cell Fact 15, 42 (2016). https://doi.org/10.1186/s12934-016-0441-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-016-0441-7